已经开发了以液相色谱串联质谱(LC-MS/MS)为基础的方法来区分糖肽的核心-和天线-岩藻糖的糖基化。通过完整的糖肽分析可以解析不同的糖基化位点(异质性)以及每个位点上可能存在的多种糖链(微观异质性)。本研究中使用含有核心-和天线-岩藻糖糖基化位点的血清糖蛋白α-1-抗胰蛋白酶(A1AT)。使用唾液酸酶去除唾液酸以简化糖基化的微观异质性,并可以增强具有类似糖链结构糖肽的MS信号。使用β1-3,4半乳糖苷酶来区分核心-和天线-岩藻糖基化。我们发现源内裂解严重影响低丰度糖链修饰的糖肽鉴定和定量。因此需要对质谱仪参数进行设定以最小化源内裂解。使用三步法质谱裂解策略进行糖肽的识别,并通过pGlyco软件注释和人工检查进一步确定。用于初始糖肽片段化的碰撞能量对提高氧鎓离子的检测以及更好地选择Y1离子(肽+GlcNAc)至关重要。结构配置显示A1AT糖肽的所有3个糖基化位点都含有复杂的N-糖链结构:Asn70位点含有没有岩藻糖基化的二天线糖链;Asn107位点含有核心-和天线-岩藻糖基化的二天线,三天线和四天线糖链;Asn271位点含有核心-和天线-岩藻糖基化的二天线和三天线糖链。Asn107位点上的核心-和天线-岩藻糖基化的相对强度与A1AT蛋白相似,表明Asn107位点的糖基化水平比其他2个位点高得多。
1.前言
异常蛋白质的糖基化尤其是岩藻糖基化与各种疾病相关,例如癌症[1]。核心岩藻糖基化是指岩藻糖连接到N-糖链的核心-N-乙酰葡糖胺上形成的,而天线岩藻糖基化则是岩藻糖连接到天线-N-乙酰葡糖胺或半乳糖上形成的。已经发现一些蛋白质的核心-或天线-岩藻糖基化的变化指示各种癌症的发生。例如,血清中α-胎儿球蛋白(AFP-L3)的核心-岩藻糖基化水平的增加被发现与肝细胞癌有关[2]。基于免疫分析法和LCA对核心-岩藻糖基化糖蛋白的高亲和力,AFP-L3使用小扁豆凝集素(LCA)印迹试验进行检测[3]。另一个例子是CA19-9,一种天线-岩藻糖基化唾液酸化路易斯A(lewis A)结构。血清中CA19-9水平的升高是胰 腺癌最广泛使用的临床指标[4]。使用唾液酸化路易斯A结构的特异性抗体通过免疫分析法来检测CA19-9[4]。该方法依赖于特异性抗体,因此不能轻易应用于其他糖蛋白。除了上述基于免疫分析法的方法之外,用于核心-和天线-岩藻糖基化分析的另一种常规方法涉及各种岩藻糖苷酶的组合以及从糖蛋白中释放糖链后通过HPLC对糖链进行几个循环的分离[5]。尽管最近开发的固定化PNGase F消化程序使糖链可以从糖蛋白中快速释放[6],但该岩藻糖苷酶消化的方法是冗长的。更重要的是,大多数蛋白质具有多个岩藻糖基化位点,上述分析丢失了位点特异性信息,因此不能提供蛋白质的核心-或天线-岩藻糖基化异常的直接证据,而该证据是精准诊断的关键。
许多研究一直在探索基于MS的完整糖肽分析,例如改善质谱仪的灵敏度,分辨率和裂解模式以及开发用于糖肽数据分析的软件[7,8]。这些研究过程中一直使用CID,ECD,ETD,EThcD,低能量和高能量HCD等裂解方式或这些裂解方式的组合来阐明糖肽的结构[9-11]。已经开发了用于阐明这些谱图的几种不同软件,其中Byonics[12]和GPQuest[13]是目前使用最广泛的软件。然而Byonics依赖于基于肽段序列的得分,低估了糖肽的假阳性发现率[14],GPQuest需要一个样品来源的肽数据库来匹配糖肽,这使得实验更加复杂[13]。在这里,我们使用了Yang PY和He SM团队中新开发的pGlyco软件来帮助糖肽的质谱分析。pGlyco使用HCD MS2产生的氧鎓离子来过滤糖肽,在Y1离子上使用HCD MS3进行肽段测序,并使用CID MS2进行糖链的解析[15]。pGlyco2.0是一个更新版本,它使用了阶梯式HCD碰撞[14]。虽然pGlyco中MS3的参与使得扫描速度稍慢,但与pGlyco 2.0相比,它能够手动检查具有更复杂片段的糖链结构和肽段序列。因此,我们使用pGlyco作为首选方法。
单独使用LC-MS/MS,由于它们在C18色谱柱上相似的保留时间和糖肽相同的质核比[16],因此通常难以区分核心-和天线-岩藻糖基化,并且在MS/MS裂解过程中岩藻糖可能发生从天线-到核心-位置的迁移[17]。糖链衍生化例如全甲基化能够解决岩藻糖迁移的问题,但是在衍生化之前需要从糖肽中释放糖链,以致位点特异性信息丢失[17,18]。因此,我们试图开发一种在进行LC-MS/MS之前区分核心-和天线-岩藻糖基化的方法,并使用pGlyco促进核心-和天线-岩藻糖基化鉴定和半定量的质谱分析。在本研究质谱分析前,我们应用唾液酸酶和半乳糖苷酶双重消化来区分核心-和天线-岩藻糖基化。使用唾液酸酶去除唾液酸以简化糖基化微观异质性并增强糖肽的MS信号。β 1-3,4半乳糖苷酶(来自牛睾丸)用于区分核心-和天线-岩藻糖基化,其中半乳糖苷酶不能从天线-岩藻糖基化的路易斯结构释放半乳糖[19]。
血清蛋白α-1-抗胰蛋白酶(A1AT)的岩藻糖基化水平已被确定为各种癌症[20,21]和炎症[22]的潜在生物标志物。本研究中,糖肽的唾液酸酶和半乳糖苷酶双重消化之后直接进行LC-MS/MS分析,而不从糖肽中释放糖链。通过对A1AT的糖肽的成功鉴定和半定量以及核心-和天线-岩藻糖基化的明确区分,解析了糖基化位点和每个位点上多种可能的糖链。
2.材料方法
2.1胰蛋白酶将蛋白质消化成肽段:我们向10μg α-1-抗胰蛋白酶(A1AT)中加入10μL 50mM的碳酸氢氨并用移液器吹打充分溶解样品。溶解的A1AT样品用10mM tris(2-羧乙基)磷化氢(TCEP)在37℃下还原30分钟并用20mM碘乙酰胺(IAA)在室温下避光烷基化15分钟。用50mM碳酸氢氨将样品溶液稀释3倍,加入1μL 0.5μg/μL的胰蛋白酶(Promega,Madison,WI)在37℃下孵育16小时。最后95℃下孵育5分钟使胰蛋白酶失活并通过真空离心机干燥样品。
2.2糖肽的富集和缓冲液置换:使用3K Ultra离心过滤器-15(Millipore Amicon)进行糖肽富集和缓冲液置换。离心机设置为7,500g,1小时,重复3次将缓冲体系由上述体系置换为25mM的乙酸钠(pH5.5)。修饰的糖肽大于3K,因此只有小于3K的非糖肽类才能透过3K滤膜。
2.3唾液酸酶/半乳糖苷酶双重消化:对于唾液酸酶和半乳糖苷酶消化,将30μL 25mM乙酸钠溶液中的糖肽混合物与15mU(3μL)来自大肠杆菌((Prozyme, Hayward, CA))中表达的非特异性α2-3,6,8,9唾液酸重组酶以及75mU(3μL)来自牛睾丸(Prozyme,Hayward,CA)的β1-3,4半乳糖苷酶在37℃下温育18小时以除去所有唾液酸酸残基和半乳糖,前提是岩藻糖没有连接到N-糖链末端的N-乙酰葡糖胺上。糖苷酶在95℃下5分钟失活。
2.4 C18脱盐:加入三氟乙酸(TFA)直到pH值达到2。用200μL含有0.1%TFA的50%乙腈溶液活化C18柱(Fisher Scientific,San Jose,CA)5次,并用0.1%的TFA水溶液平衡3次,每次1500g/min离心1min。将多肽与C18珠子结合5次,然后用0.1%的TFA洗涤3次通过上述离心程序去除非特异性结合;使用20μL含有0.1%TFA的50%乙腈溶液通过上述离心程序进行洗脱。重复洗脱一次,然后通过真空离心机干燥合并的洗脱液。
2.5 糖肽的LC-MS鉴定
Nano LC-MS/MS工作条件如先前研究中所述[23]。C18毛细管柱(100μm×15cm;3μm粒径,200?))(Thermo Fisher Scientific,SanJose,CA)用于LC分离,梯度洗脱使用Ultimate 3000 nanoLC系统(Thermo Fisher Scientific,SanJose,CA),流速为350nL/min。流动相A为0.1%的甲酸水溶液中含有2%的乙腈,流动相B为乙腈溶液中含有0.1%的甲酸和2%的水。分析方法的梯度时长100分钟,其中平衡10分钟后,流动相B的梯度组成为:2min内由3%上升到7%,8min内再从7%上升到14%,55min内从14%上升到25%随后进行洗涤和平衡步骤:其中流动相B在5min内增加至90%并保持8min,然后在0.1min内返回至3%并保持17min。
使用以正离子模式操作的Orbitrap Fusion Lumos质谱仪(赛默飞世尔科技公司,圣荷西,美国加州)进行分析。ESI喷雾电压和毛细管电压的设定如下文所述。对每个样品进行两次LC-MS分析。每次运行包括两个连续的MS扫描类型。在第一次运行中,通过低能量HCD MS2检测氧鎓离子138.05来选择糖肽;来自糖肽的Y1离子(肽+GlcNAc)通过高能量HCD MS3用于肽段测序。在第二次运行中,在通过低能HCD MS2选择糖肽后,将所选择的糖肽进行CID MS2用于糖链的结构分析。正如结果部分所讨论的那样,为了更好地检测氧鎓离子,更好地选择Y1离子以及更好地裂解Y1离子和糖链,每个裂解步骤的碰撞能量也进行了优化。全扫描定义质量范围为m/z 600到1800,MS/MS采用速度模式。
2.6 糖肽鉴定的数据库检索:
He SM团队开发的搜索引擎pGlyco和pFind用于糖蛋白的分析。两次LC-MS运行的原始数据首先需要一致以确保两次运行中相同前体离子的保留时间相同。pFind通过使用来自第一次LC-MS运行的MS3谱图来进行Y1肽段鉴定:(1)固定化修饰:半胱氨酸的脲基甲基化(+57.021Da);(2)动态修饰:甲硫氨酸氧化(+15.995Da)和N-乙酰己糖胺(+203.075Da);(3)允许错过一次裂解;(4)肽段离子公差:15ppm;(5)碎片离子公差:25ppm。从第一次LC-MS运行中鉴定的Y1肽段和第二次LC-MS运行的原始数据被输入到pGlyco中进行糖肽匹配和评分。所有鉴定的糖肽均由EURO CarbDB开发的GlycoWorkbench软件进行手动检查[24]。根据糖生物学基础[25]使用糖链的命名规则,根据NIBRT GlycoBase使用缩写。
3.结果与讨论
N-连接糖蛋白的两种岩藻糖基化结构,核心-和天线-岩藻糖基化,已被认为是各种癌症中的生物标志物[1]。通常很难区分这两种结构。因此,我们试图开发一种方法来区分目标蛋白的每个糖基化位点上的核心-和天线-岩藻糖基化。本研究的工作流程如图1所示。简而言之,将血清蛋白α-1抗胰蛋白酶(A1AT)标准品消化成肽段,然后通过唾液酸酶/半乳糖苷酶双重消化处理用于糖基的切除。将处理后的糖肽半富集并通过3K滤膜脱盐后直接通过LC-MS/MS进行分析。因此A1AT的核心-和天线-岩藻糖基化被成功地区分和定量。
血清蛋白的总体岩藻糖基化水平相当低[26]。利用用于肽分析的常规质谱设置,将发生糖肽的源内碰撞诱导的解离(也称为喷雾分离器解离,在此缩写为源内解离)。这是离子从大气压离子源转移到质谱仪的真空室期间由于碰撞激发而导致解离的过程[27]。在我们的实验中,发现糖肽的源内解离严重影响低丰度岩藻糖基化肽段的鉴定和半定量。但是,迄今为止还没有对这个问题进行详细分析。以A1AT糖肽中最丰富的糖基修饰类型A2为例,A2在唾液酸酶/半乳糖苷酶双重消化后应含有3个Hex(甘露糖),4个HexNAc(N-乙酰葡糖胺),0个NeuAc,0个NeuGlc和0个dHex,在这里简写为34000。如图2所示,使用赛默飞开发的Orbitrap Fusion质谱仪(离子传输管温度=300℃,RF=30%)[28],通过“常规”方法设定或更严格的设定进行肽段分析发现超过20%的34000糖链被衰减到23000(以XIC23000 / XIC34000计算)。因此优化了用于糖肽分析的一系列MS设定。我们发现在较低温度和较低的RF(150℃,20%)条件下,糖肽的源内解离降低到3%以下,而含有34001糖链的核心-岩藻糖基化肽段的信号没有显著降低。我们还发现,较高的喷雾电压(喷雾电压> 2300 V)会提供更高的信号,但同时也增加了源内解离。因此使用稳定喷雾的喷雾电压2300 V。该优化的设定条件用于进一步鉴定A1AT糖肽的位点特异性糖基化以及对核心-和天线-岩藻糖基化进行半定量。
图3显示了直接的LC-MS/MS策略,首先通过用低能HCD MS2检测氧鎓离子138.05来选择糖肽(图3A);然后将来自糖肽片段的Y1离子进行高能量HCD MS3(图3B)用于肽段测序;而所选择的糖肽进行CID MS2(图3C)用于糖链的结构分析;图3D中总结了整个流程。第一步的HCD碰撞能量(CE)对糖肽的片段化至关重要。它针对提高氧鎓离子的检测和Y1离子的选择进行了优化。如图4所示,对于含有Asn271位点的非岩藻糖基化以及核心-岩藻糖基化或天线-岩藻糖基化的二天线糖肽,当CE为24(从CE20到CE32的一系列HCD中)时的低能量HCD MS2提供了最强的Y1离子片段。这种最适的CE似乎与糖链结构或肽段序列无关。如糖肽(含有Asn271位点)所示,无论是非岩藻糖基化还是核心-岩藻糖基化或天线-岩藻糖基化结构都具有相同的最适CE(图4),其中AlAT的其他2种糖肽(含有Asn107或Asn70位点)也具有相同的最适CE(补充信息-图S1)。
与此相反,第二步用于肽注释的HCD CE和第三步用于糖链注释的CID CE灵敏度却不高。当CE为35、38或40时,HCD MS3均显示出相似的裂解模式(如图5所示),其中HCD MS3 CE35对较高m/z的片段产生的信号稍强。糖肽的糖链结构在CID CE30和CE35下均显示出相似的裂解模式(图6)。因此在随后的实验中,分别使用低能量HCD CE24,CID CE30和高能量HCD CE 35。
唾液酸酶释放α 2-3,6,8,9-N-乙酰神经氨酸,导致N-糖链的末端变为半乳糖。随后,β-半乳糖苷酶切除β 1-3,4半乳糖,前提是没有岩藻糖与N-糖链末端N-乙酰葡糖胺结合,因此提供了区分核心-岩藻糖基化和天线-岩藻糖基化的方法[19]。在唾液酸酶消化的样品中核心-岩藻糖基化和天线-岩藻糖基化糖肽具有相同的m/z(图7.B),因此唾液酸酶消化样品的图谱是核心-和天线-岩藻糖基化肽段的混合物(图7.B1)。相比之下,唾液酸酶/半乳糖苷酶双重消化的样品中核心-和天线-岩藻糖基化肽段具有不同的m/z(图7.A)。因此,使用唾液酸酶/半乳糖苷酶对糖肽进行双重消化后,不需要进一步的MS/MS分析或大量连续的外切糖苷酶消化就可以区分出两种类型的岩藻糖基化,这与之前在糖链水平上的工作相似[19]。此外,用唾液酸酶/半乳糖苷酶双重消化后,天线-岩藻糖基化糖肽的保留时间早于其相应的核心-岩藻糖基化糖肽(图7.A),表明半乳糖增强了其亲水性。我们发现核心-和天线-岩藻糖基化糖肽的洗脱时间和裂解模式都不相同。在核心-岩藻糖基化糖肽的CID MS/MS图谱中,可以观察到几个核心-岩藻糖基化糖肽片段(图7.A1);而在天线-岩藻糖基化糖肽的CID MS/MS图谱中出现的三个信号最强的片段为pep-43001,pep-43000和pep-33001(图7.A2)。其他糖肽中也发现了核心-和天线-岩藻糖基化裂解模式的差异(图S2)。因此,使用唾液酸酶/半乳糖苷酶双重消化后,可以通过直接LC-MS/MS分析获得核心-和天线-岩藻糖基化肽段之间的稳定差异。传统的外切糖苷酶与岩藻糖苷酶α 1-2,3,4,6和岩藻糖苷酶α 1-3,4也可以进一步应用于唾液酸酶/半乳糖苷酶双重消化的糖肽,显示了该策略的有效性(图S3)。
在之前的研究中,我们分析了从糖蛋白A1AT上释放的糖链,发现A1AT在唾液酸酶/半乳糖苷酶双重消化后有12种不同的糖链结构[19]。在本研究中,我们鉴定了10个特定糖基化位点上的这些结构。它们在C18色谱柱上的保留时间及其相对强度如表1所示。在补充信息-图S2中显示了所有鉴定糖肽的低能量HCD MS2,高能量MS3和CID MS2图谱。含有Asn107位点的糖肽1 ADTHDEILEGLNFnLTEIPEAQIHEGFQELLR(修饰的氨基酸显示为小写)具有最多种类的糖链修饰类型,包括A2,FA2,A2FG,A3,FA3,A3FG,A4,FA4,A4FG,A3F2G2,而其他两个位点具有比较少的糖链修饰类型(含有Asn271位点的糖肽2YLGnATAIFFLPDEGK:A2,FA2,A2FG,A3,A3FG和含有Asn70位点的糖肽3 QLAHQSnSTNIFFSPVSIATAFAMLSLGTK:A2)。每个位点的糖基化程度可能取决于蛋白质结构或者位点与某些氨基酸或N/C末端的接近程度[22]。如图8(晶体结构来自[29])所示,所有三个位点都位于蛋白质表面和环中,其中Asn107几乎位于大环的中心,并且可能更易接近各种糖基转移酶,而Asn271和Asn70更接近α螺旋或β折叠结构并且空间较小。这可能在某些方面解释了为什么Asn107具有最多的糖基化修饰类型。
正如预期所示,在唾液酸酶/半乳糖苷酶双重消化后,核心-岩藻糖基化糖肽额外的岩藻糖使得糖肽更具亲水性,因此其从C18柱的洗脱早于其相应的二天线-和三天线糖链修饰的非岩藻糖基化糖肽。与相应的核心-岩藻糖基化的情况相比,额外的半乳糖使天线-岩藻糖基化的二天线糖肽更加亲水。然而,进一步的半乳糖和/或岩藻糖没有使得三-或四-天线糖肽更加亲水,并且所有岩藻糖基化的三天线糖肽或四天线糖肽在非常缓慢的洗脱梯度(0.2%ACN /min)下洗脱时间相似。这表明在唾液酸酶/半乳糖苷酶双重消化后,糖肽的疏水性主要由其肽骨架决定,只在二天线糖链中发现有轻微的疏水性变化。这种变化可能是由于与三天线-和四天线-糖链相比,二天线糖链具有较少的糖基,因此一个或两个额外的糖基就可以导致糖肽总体疏水性更高。
我们先前对糖链的研究表明,非岩藻糖基化的二天线-和三天线-糖链是A1AT中最丰富的两个糖链结构,分别占35.8%和25.2%[19]。在糖肽1和糖肽2的研究中,我们发现Asn107上的二天线-和三天线-糖链修饰分别占48%和34%,而在Asn271上分别为97%和1%(表1)。糖肽3的分析显示Asn70上仅有A2糖链修饰;因此我们认为Asn70上的糖链修饰对A1AT蛋白的整体糖基化没有多少贡献。卡方检验用于比较从A1AT蛋白质释放的糖链类型(数据来源于以前的结果[19])和糖肽1(含有Asn107位点)的糖链修饰类型或糖肽2(含有Asn271位点)的糖链修饰类型(表2)之间的主要糖链修饰类型的相对峰强度。从A1AT蛋白释放的糖链类型与Asn271上的糖链修饰类型显著不同(p值一项经典的凝集素印迹试验研究发现,核心-岩藻糖基化而非天线-岩藻糖基化的A1AT的上调可以作为肝细胞癌诊断的指标[20],而A1AT的天线-岩藻糖基化可以作为炎症发生的指标,尤其是在HBV感染的患者中[22]。我们先前的糖链研究表明,二天线-核心-岩藻糖基化是A1AT蛋白质中最丰富的核心-岩藻糖基化类型。从表1中我们可以推断,如果患者的A1AT核心-岩藻糖基化类型发生改变,Asn107位点上的二天线-核心-岩藻糖基化很可能是通过质谱法进行精确监测和定量的合适目标。另一种经典的基于凝集素印迹的研究表明,A1AT岩藻糖基化水平的总体升高能够区分肺腺癌与良性病变或其他肺癌亚型[21]。本研究中开发的策略将能够鉴定和定量A1AT特定位点上的核心-和天线-岩藻糖基化。在今后的工作中,这种方法将被用于研究各种癌症发展过程中血清A1AT糖基化的变化。包括特异性位点信息的更精确的岩藻糖基化分析能够提供更高的诊断价值。此外,该策略可应用于各种疾病发展期间其他关键糖蛋白的研究。
4.结论
我们已经开发了一条渠道来研究A1AT的糖基化,以确定核心-与天线-岩藻糖基化的存在,而不用从糖肽中分离糖链和肽段。这是在A1AT蛋白质标准品上进行的,该蛋白质经胰蛋白酶酶切,随后进行唾液酸酶/半乳糖苷酶双重消化。除了末端N-乙酰葡糖胺被岩藻糖基化修饰外,半乳糖苷酶可以去除N-糖链中的末端半乳糖残基,从而提供区分核心-岩藻糖基化和天线-岩藻糖基化的手段。糖链的位点和结构可以通过该方法同时鉴定。我们在糖肽QLAHQSnSTNIFFSPVSIATA的Asn70位点上鉴定了1个糖链结构(A2),在糖肽ADTHDEILEGLNFnLTEIPEAQIHEGFQELLR的Asn107位点上鉴定获得10个糖链结构(A2,FA2,A2FG,A3,FA3,A3FG,A4,FA4,A4FG和A3F2G2)以及糖肽YLGnATAIFFLPDEGK的Asn271位点上鉴定获得5个糖链结构(A2,FA2,A2FG,A3和A3FG)。我们相信该方法将广泛用于在各种疾病发展期间对A1AT或其他关键糖蛋白上的核心-和天线-岩藻糖基化进行鉴定和定量。
补充信息
图S1糖肽1,糖肽2或糖肽3具有相同的最适低能量HCD CE24。
图S2所有鉴定的具有不同糖基修饰类型糖肽:糖肽1(含有Asn107位点),糖肽2(含有Asn271位点)和糖肽3(含有Asn70位点)的低能量HCD MS2,高能量HCD MS3和CID MS2图谱。
图S3唾液酸酶/半乳糖苷酶消化的A1AT样品,唾液酸酶/半乳糖苷酶/α 1-2,3,4,6岩藻糖苷酶消化的A1AT样品以及唾液酸酶/半乳糖苷酶/α 1-3,4岩藻糖苷酶消化的A1AT样品中的岩藻糖基化糖肽的XIC(提取离子流图)图谱。
表1.双重消化后的A1AT糖肽以及每个位点的核心-和天线-岩藻糖基化的相对峰面积的总结。
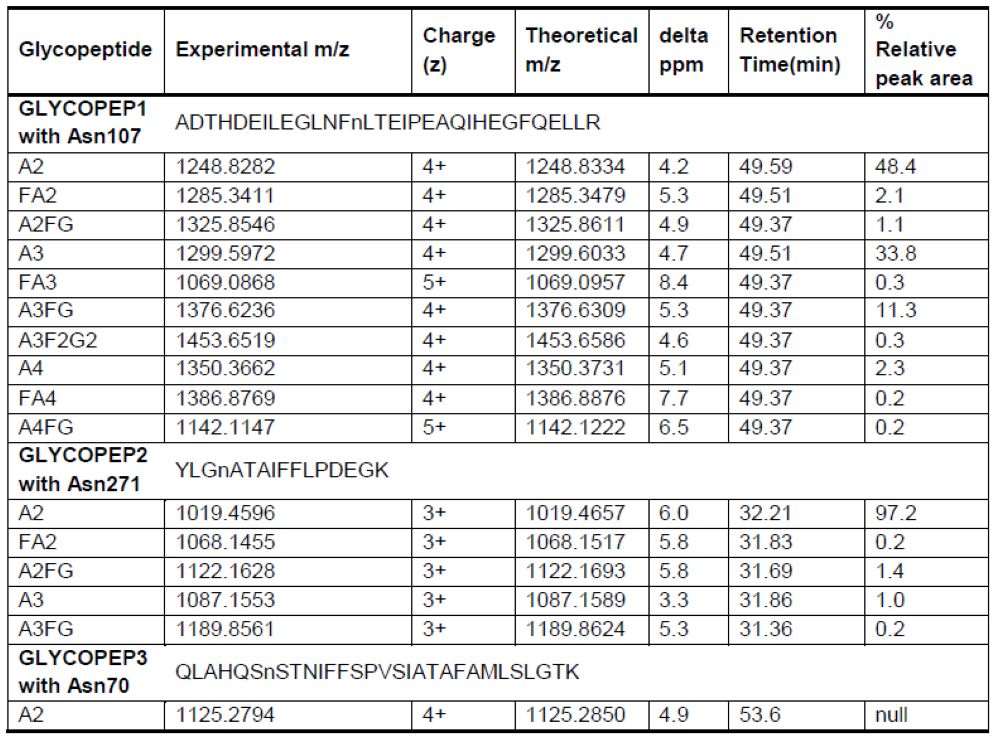
表2.卡方检验用于比较从A1AT蛋白质释放的糖链类型(数据来源于以前的结果[19])和糖肽1(含有Asn107位点)的糖链修饰类型或糖肽2(含有Asn271位点)的糖链修饰类型(表2)之间的主要糖链修饰类型的相对峰强度。
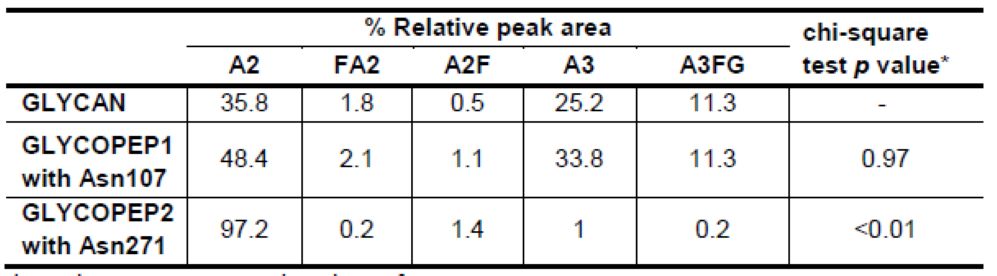
* p值< 0.05 认为差异显著

图1 鉴定A1AT糖基化的实验工作流程。首先将糖基化的A1AT酶切成肽段,然后通过唾液酸酶/半乳糖苷酶双重消化进行糖基的切除。将糖肽进行直接LC-MS/MS分析而不需要释放糖链。
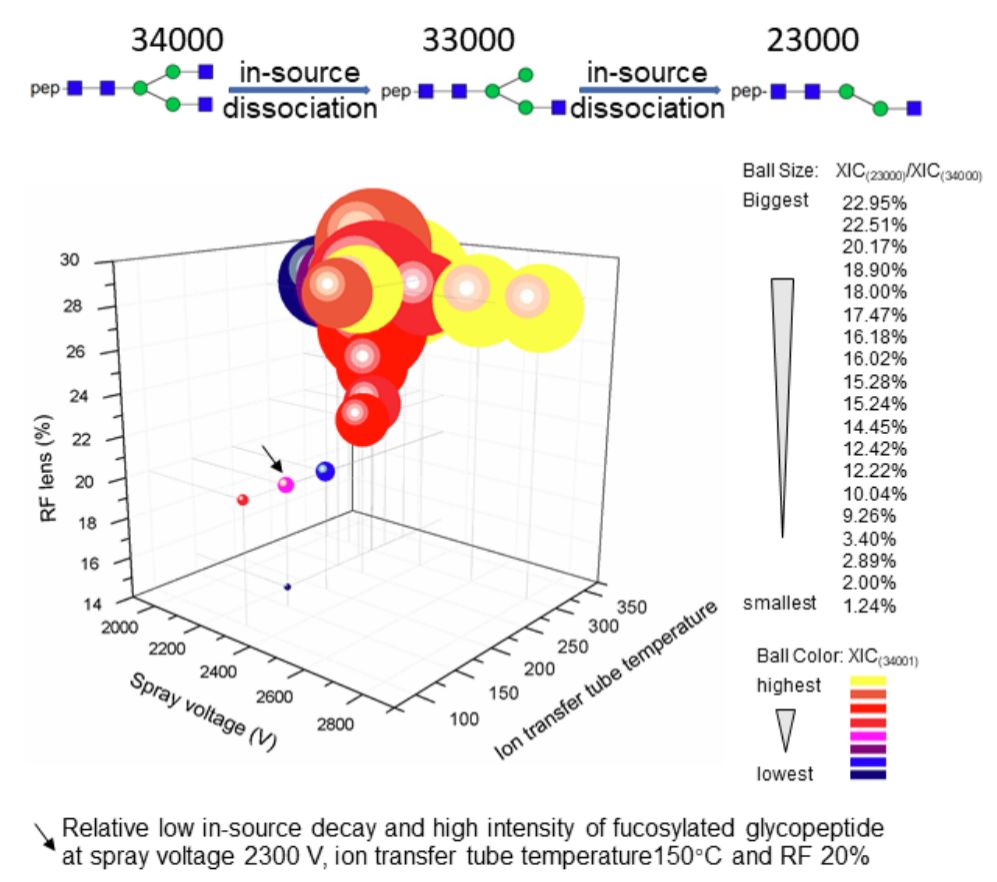
图2在不同的离子传输管温度,喷雾电压和RF%设定条件下,具有A2和FA2糖基修饰类型的A1AT糖肽(Asn271)的源内裂解。

图3具有A2FG糖基修饰类型的A1AT糖肽(Asn271)的图谱:(A)低能量HCD MS2图谱;(B)低能量HCD诱导的高能量HCD MS3图谱;(C)低能量HCD诱导的CID MS2;(D)糖肽裂解的示意图。
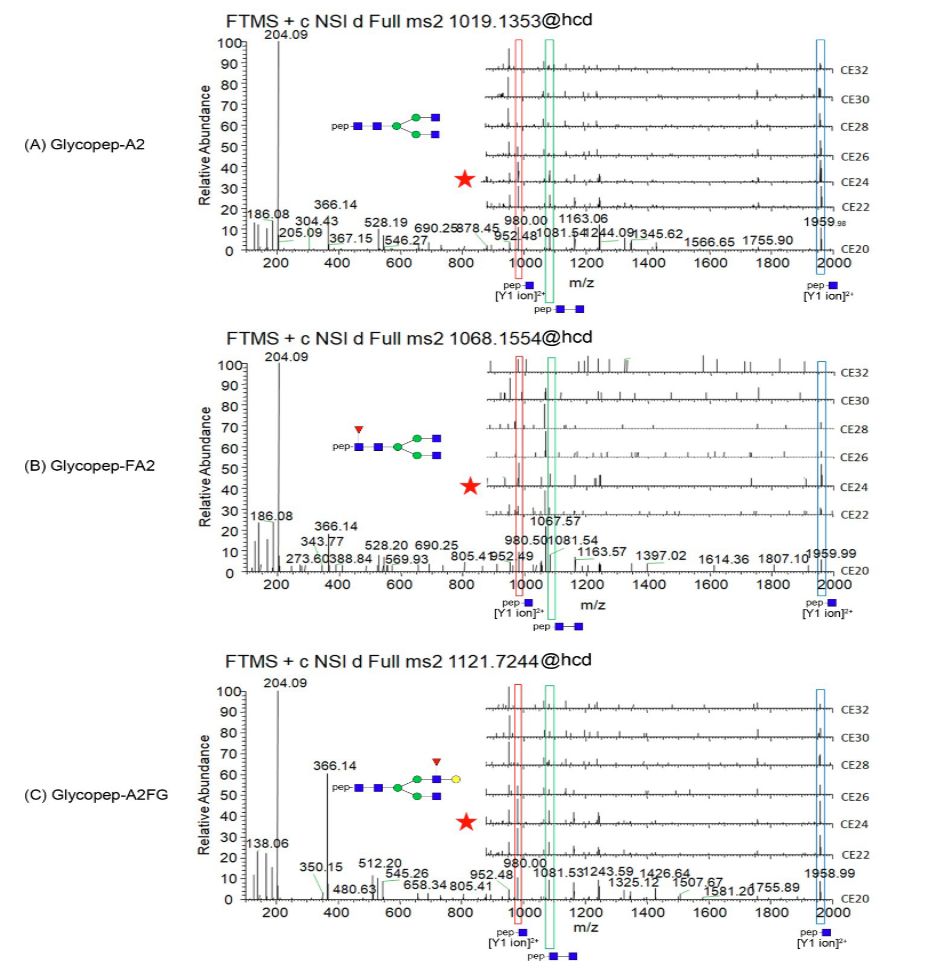
图4 各种低能量HCD碰撞能量下具有A2,FA2和A2FG糖基修饰类型糖肽(Asn 271)的MS2裂解模式,表明CE24下的低能量HCD产生信号最强的Y1离子(肽+GlcNAc)。
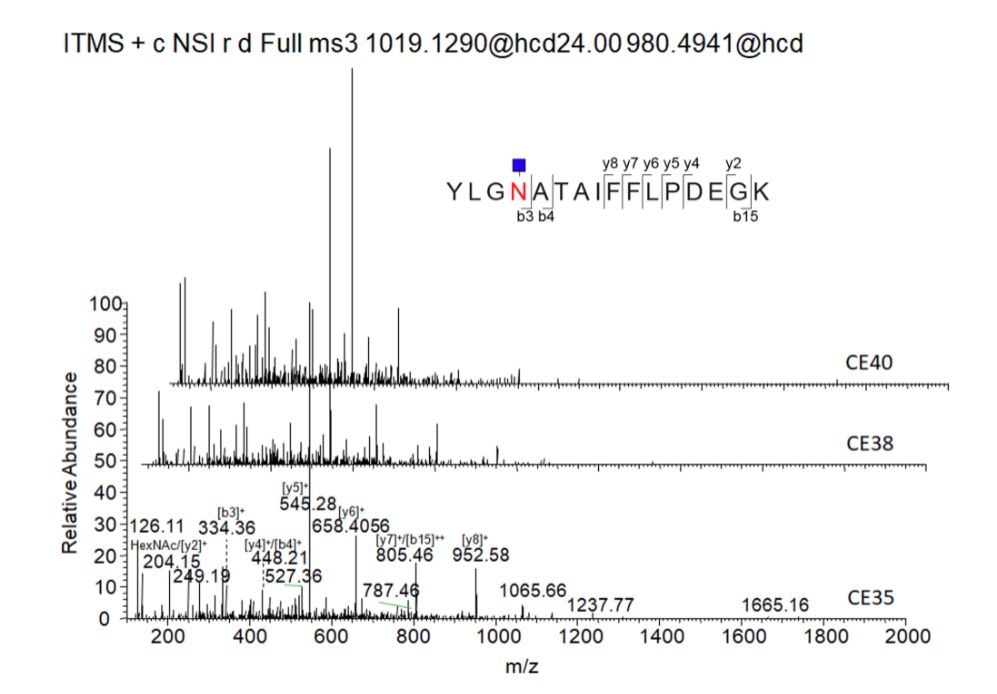
图5 各种高能量HCD碰撞能量下糖肽(Asn271)Y1离子(肽+GlcNAc)的MS3裂解模式,表明在CE35下的高能量HCD提供了的裂解图谱。
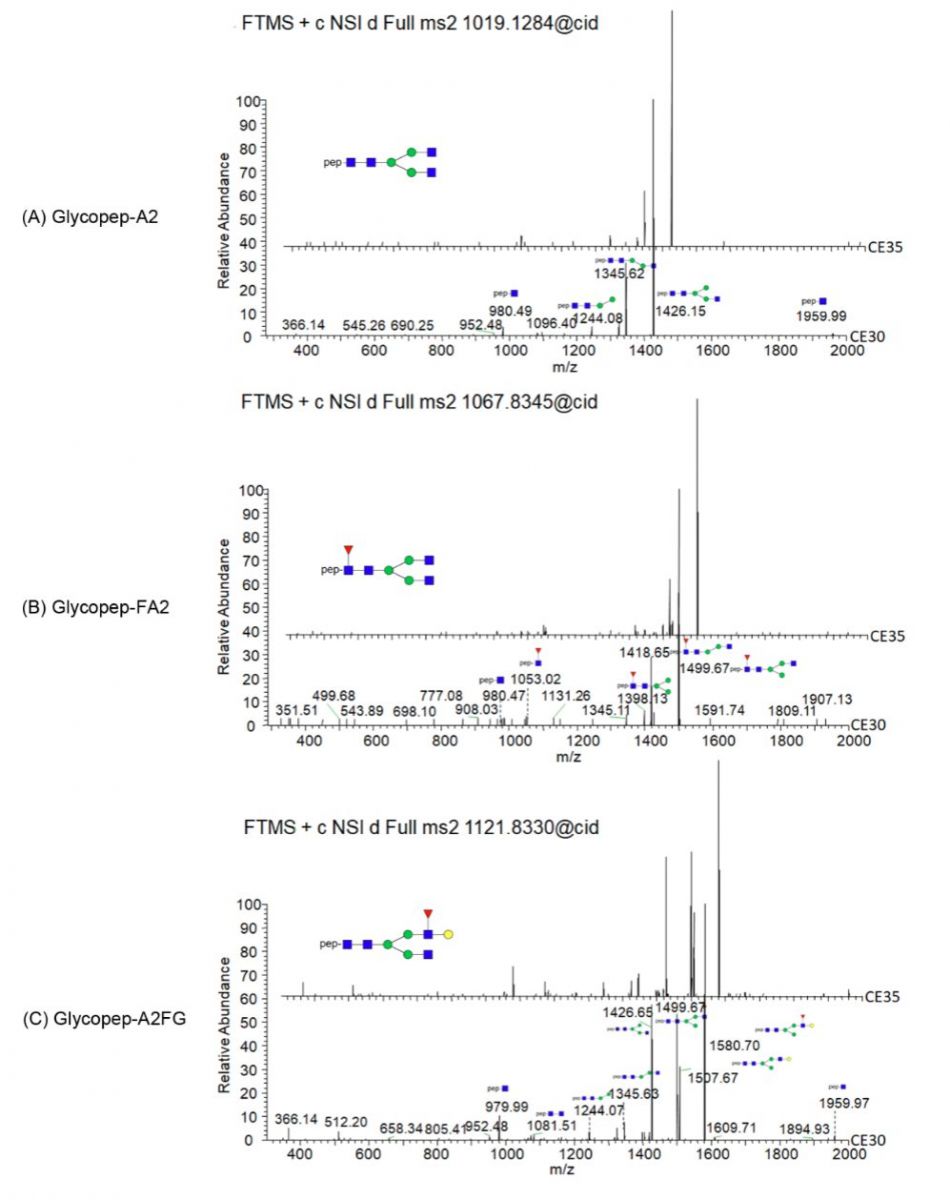
图6 在各种CID碰撞能量下具有A2,FA2和A2FG糖基修饰类型的糖肽(Asn271)的MS2裂解模式,表明在CE30或CE35下这些糖肽获得了相似的裂解图谱。

图7 通过唾液酸酶/半乳糖苷酶消化来区分糖肽(Asn271)的核心-和天线-岩藻糖基化(A:保留时间;A.1:FA2的图谱;A.2:A2FG的图谱)。作为对照,唾液酸酶消化的情况下没有观察到差异(B:保留时间;B.1:A2G2(F)的图谱)。
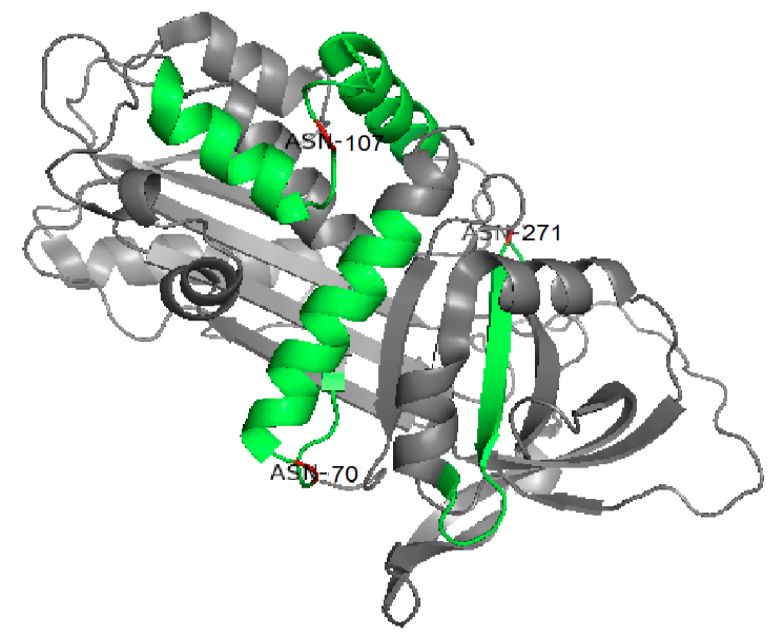
图8 在3D结构中A1AT的三个糖基化位点Asn70,Asn107和Asn271标记为红色,通过质谱检测到的肽段标记为绿色。





合作咨询
![]() 肖女士
肖女士
![]() 021-33392297
021-33392297
![]() Kelly.Xiao@imsinoexpo.com
Kelly.Xiao@imsinoexpo.com
 2006-2024 上海博华国际展览有限公司版权所有(保留一切权利)
沪ICP备05034851号-57
2006-2024 上海博华国际展览有限公司版权所有(保留一切权利)
沪ICP备05034851号-57