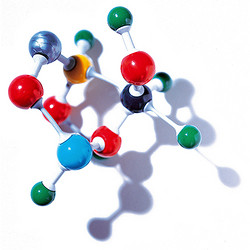
第二部分:APOE基因变异,变了一个胺基酸,变了命运
3.APOE基因型与相应的分子结构和功能
APOE基因有三种常见的变异型,分别是E2, E3和E4变异型,互相之间,仅有一个核苷酸的区别。E3型最常见,约占65-70%的比例,而E4型约占15-20%,E2型占约5-10%。E3型被认为是正常型,以E3型为参照,E4型显著增加老年痴呆症风险,而E2型微小减弱风险。我们先从分子生物学和蛋白质结构的角度来看发生了什么?apoE蛋白分子上有两个重要功能区域,受体结合区域(LDL receptor binding domain)和油脂结合区域(lipid binding domain)。受体结合区域位于蛋白质中部,富含带正电荷的碱性胺基酸,有利于与带负电荷的细胞表面受体结合。油脂结合区域位于蛋白质尾部(C-端), 富含疏水胺基酸。APOE基因的E4和E2变异型表现为两个位置上的胺基酸改变:E4变异使第130号位置从半胱胺酸Cys变为精胺酸Arg,而E2变异使第176号位置从精胺酸Arg变为半胱胺酸Cys。也就是说正常的E3型蛋白质apoE3是130Cys,176Arg;高风险E4型蛋白质apoE4是130Arg, 176Arg;而低风险E2型蛋白质apoE2是130Cys, 176Cy。这些胺基酸改变会造成什么样的功能变化呢?
第一,简单的从胺基酸层面看,我们不难看出,apoE4在130号位置增加了一个正电荷胺基酸Arg,而apoE2在176号位置损失了一个正电荷胺基酸Arg。有趣的是,第130和176号位置分别位于apoE蛋白质的受体结合区域的两侧。apoE受体结合核心区域位于152-168号位置,这个区域富含带正电荷的碱性胺基酸,用于与富含负电荷胺基酸的受体结合。apoE2蛋白质在靠近受体结合区域位置上失去了一个正电荷胺基酸。实验证明apoE2与其在肝细胞上的受体LDLR的结合能力却实大大下降,只有正常的apoE3的2%。而apoE4蛋白质在距受体结合区域稍远的位置上增加一个带正电荷的胺基酸,apoE4其与受体的结合能力则比正常的apoE3微小增加。因此,如果用与受体的结合能力来评价功能改变,apoE3为正常型,apoE2为严重失活型,而apoE4为微弱增活型。在人群中我们则观察到,携带有E2/E2纯合子变异的人容易患一种称为三型高血脂的疾病,此病的重要特症是血液中乳糜微粒和其他脂蛋白残体不能被肝细胞清除,所以也称为残体代谢病(remnant metabolic disorder)。更进一步的观察还发现,APOE基因上其他几个在受体结合区域失去带正电荷基酸的变异,也都会导致家族性三型高血脂症,进一步肯定了这一区域失去正电荷会系统性的损坏apoE与其受体的结合能力。apoE与受体的结合能力是apoE4>apoE3>>apoE2。仅从受体结合能力的角度,我们很难解释为什么只微弱增加受体结合能力的E4变异会那么显著的增加老年痴呆症风险,而受体结合能力损失了90%的E2变异确只微弱减轻风险。因此我们推断,apoE与受体结合的通道可能不是其主要的老年痴呆症致病机制。
第二,从蛋白质三维结构的变化角度我们可以看到,E4变异会引起apoE4蛋白质区域间出现新的盐键。在130号位置引入一个带正电荷的胺基酸Arg, 会导致一个特殊的新盐键,这个盐键恰好锁住了apoE蛋白质尾部的脂质结合区域,使这一区域丧失了灵动性,进而影响了这一区域与油脂分子的结合能力和选择性。见示意图。逐一置换APOE基因上胺基酸的实验证明,仅有E4变异所在的位置才可能通过盐键改变脂质结合区域的构像。这暗示E4变异造成脂质结合区构象的改变是唯一的和特殊的,其它位置上的胺基酸变化几乎没有可能获得类似的构象变化。在现实生活中,确实没有发APOE基因上的其他突变增加老年痴呆症风险。
现在我们来看看因E4变异而锁住尾部脂质结合区域可能导致什么样的功能变化。实验数据显示,apoE4脂蛋白微粒的含脂量比正常的apoE3少2-3倍。而apoE2虽然与受体的结合能力大为下降,但其含脂量不受影响,与正常的apoE3相当。除了含脂量的多少外,apoE4更喜欢与甘油三酯亲和,而与胆固醇及磷脂的亲和力下降。apoE2和apoE3能更好的与胆固醇和磷脂亲和。从脂蛋白含脂量的角度来看看,apoE4<
4.APOE与淀粉样蛋白斑块形成假说
APOE是老年痴呆症重中之重的风险基因, 有很多假说试图解释APOE如何导致老年痴呆症,这些假说复盖了几乎所有可能的病因通道。这里我们只作重讨论APOE可能通过什么样的机理导致粉状蛋斑块沉积。我们知道携带APOE基因E4变异的患者不但发病时间早,脑中的淀粉样蛋白沉积更多,斑块的密度也更高。可以猜测影响大脑中淀粉样蛋白斑块形成的途径主要有三个:过度产生,清除受阻,聚合加速。因为apoE是一个脂蛋白,并不直接参与淀粉样蛋白的产生,那么扰乱清除和加速聚合沉积可能是最主要的机制。我们先来看看正常生理状态下大脑中淀粉样蛋白是如何产生和清除的。
淀粉样蛋白的生理功能尚不清楚。淀粉样蛋白主要由神经细胞产生,神经细胞中的淀粉样蛋白前体APP经由几个酶的分步切割而产生一组长度为30-51个胺基酸的短肽并释放到细胞外,这些短肽称为淀粉样蛋白β多肽(amyloid β peptide, or Aβ)。最常见的Aβ短肽有两种,Aβ40和Aβ42。Aβ40含40个胺基酸,是神经细胞正常情况下主要产生的一种,这种短肽的水溶性更好,不易发生自身聚合。Aβ42含42个胺基酸,比Aβ40在C-端多了两个胺基酸,远比Aβ40更容易发生聚合。分析老年痴呆症病人脑中的斑块发现,有的斑块全部由Aβ42组成,有的斑块则以Aβ42为主伴有少量的Aβ40。大多数Aβ短肽分子在细胞外间质中的构像是可溶性的球状结构,称为可溶性Aβ。但在一定条件下,Aβ短肽,特别是Aβ42之间可以通过氢键反相聚合成形成有序的纤维状寡聚体,再进一步聚合成β片层结构(β-sheet)。这些非常有序的片层结构聚合物会越聚越大,因为疏水基团暴露于其表面而难溶于水,结果在细胞外间质中形成看起来像淀粉一样的沉淀物,这些沉淀物堆聚而成的斑块称为淀粉样蛋白斑块(amyloid β plaques)。这里β是指通过β片层结构形成的淀粉样蛋白斑块。详见示意图示。
绝大部分可溶性Aβ是通过血脑屏障转运到血液中而被清除的。在血脑屏障上表达的脂蛋受体LRP1 被认为是负责转运可溶性Aβ进入外周血液的主要通道。小鼠同位素标记灌注实验显示,LRP1转运可溶性Aβ的速度非常快,并且转运Aβ40的速度比是转运Aβ42的速度快约两倍。但是,如果有apoE同时存在,无论是高度脂质化的apoE还是裸体apoE, 都会使可溶性Aβ的转运速度大为下降。我们前面讲过,LRP1也是apoE的受体,以上实验结果似乎暗示,一旦apoE与LRP1结合,就会阻碍可溶性Aβ通过血脑屏障清除。我们前面讲过,不同变异型apoE与受体的结合的能力是APOE4>APOE3>>APOE2,那么可以猜测,apoE与受体的结合能力越强,其阻碍可溶性Aβ通过LRP1通道清除出大脑的能力就越强,反之亦然。支持apoE阻碍可溶性Aβ清除的证据还来自小鼠基因剔出实验(APOE knockout mouse)。在老年痴呆症模型小鼠上剔除APOE基因,脑中的淀粉样蛋白斑块会显著减小。除了LRP1通道外,另一个在血脑屏障上表达的脂蛋白受体LRP2 (LDL receptor related protein2) ,可以转运与apoJ脂蛋白微粒结合的不溶性Aβ。LRP2基因及编码apoJ的CLU基因都是老年痴呆症风险基因。除了通过血脑屏障清除A?外,还有一部份Aβ,特别是不溶性Aβ,可能是通过与apoE脂蛋白结合,然后通过与细胞表面的脂蛋白受体结合转运进入细胞中,并在细胞内的溶酶体中被降解掉。大脑中的星型胶质细胞(astrocytes), 小胶质细胞(microglia)和神经细胞上都表达LDLR和LRP1, 可以与apoE结合。另外,小胶质细胞作为大脑中的免疫细胞,还可以通过非特异性内吞机制(phagocytosis)把淀粉样蛋白斑块内吞到细胞中加以消化。与内吞机制相关的基因ABCA7和TREM2基因也与老年痴呆症风险強相关。
前面我们分析了E4变异对蛋白质三维构象的改变。E4变异严重改变了蛋白质尾部脂质结合区域的灵活性,这一结构变化使apoE4与极性的胆固醇分子和磷脂结合较差,导致apoE4脂蛋白微粒含脂量显著下降。我们可以想象星型胶质细胞合成apoE脂蛋白微粒并分泌到细胞外,与神经细胞分泌到细胞外的Aβ短肽混合共处于细胞间质,apoE脂蛋白上的油脂分子,特别是胆固醇,可能会与Aβ短肽上暴露于表面的疏水性胺基酸亲合,从而打破Aβ短肽自身聚合的平衡点,影响淀粉样蛋白的形成。就象结晶过程中当溶液浓度达到临界点时,还需要有晶种才能触发结晶一样。如果apoE脂蛋白上的胆固醇分子可以干扰Aβ聚合”晶核”形成,那么即便脂蛋白上微小变化就有可能显著改变Aβ聚合最终产量。让我们先回顾一下, apoE主要由星型胶质细胞合成, 与同样在星型胶质细胞中合成的胆固醇和磷脂等油性分子一起形成HDL like脂蛋白微粒,并分泌到细胞外,其主要生理功能被认为是为神经细胞运送胆固醇和磷脂,用于神经细胞的维修和扩增。胆固醇和磷脂都是极性油脂,就像洗涤剂一样,可以乳化融合含有疏水基团的分子。不难想象,apoE脂蛋白含脂量越高,乳化融合Aβ的容量越大,反之亦然。这样,我们就可以解释为什么apoE4因为含脂量低,兼融Aβ的容量小,从而化解Aβ聚合“晶核“形成的能力就差。因此,Aβ更易在携带E4变异人群的大脑中聚合成淀粉样蛋白斑块。实验证据显示,在饱和浓度的Aβ溶液中,加入脂质化的apoE会大大推迟Aβ自身聚合反应发生的时间。这里,apoE脂蛋白的作用是减慢聚合核的形成。更多实验显示,胆固醇分子而不是磷脂分子,可以有效阻碍胰 腺中一种叫做Amylin 的淀粉样蛋白聚合。并且胆固醇分子的前体分子lanosterol可以有效溶解眼睛白内障中不溶的蛋白质。包括Aβ淀粉样蛋白在内,这几种不溶性蛋白质都是β片层结构形成的聚合物。
支持apoE含脂量假说的一个重要证据来自ABCA1基因。apoE的脂质化程度主要受ABCA1基因调节。ABCA1是一种分子泵,其功能是将细胞中胆固醇导流到含有apoE或apoJ的HDL-like脂蛋白微粒上。激活ABCA1表达可以显著增加apoE脂蛋白微粒的含脂量。ABCA1基因的表达受视黄酸受体(retinoid receptor X, RXR)调节。视黄酸是维生素A衍生物,化学合成的RXR拮抗剂以及活化的维生素A都能有效激活ABCA1基因表达。给老年痴呆症模型老鼠喂食RXR激动剂的实验发现,小鼠脑中淀粉样蛋白斑块会在喂食RXR拮抗剂几小时后开始大量清除。并且,ABCA1趋动的淀粉样蛋白清除依赖于APOE基因的表达,在APOE基因剔出的小鼠中不起作用。为什么这么有效的RXR激动剂不用来治疗人的老年痴呆症呢?可惜将RXR激动剂用于人体临床实验时,虽然也观察到淀粉样蛋白斑块清除,但是该激动剂在很短的用药时间里即造成病人血脂极度升高,不得终停止这项人体临床试验。其他几个能活化ABCA1表达的激动剂药物,在老年痴呆症小鼠模型中也得到同样的结果。ABCA1基因突变与老年痴呆症密切相关,己经发现这个基因的失活变异增加老年痴呆症风险2-3倍,而ABCA1上的增活变异则能降低风险。
除了以上APOE淀粉样蛋白假说外,有不少实验工作显示,APOE基因E4变异增加大脑中负责免疫功能的胶质小细胞的活性和炎症反应。另一些证据则指向,表达E4的神经细胞会生产更多容易聚合的Aβ42。并且带有E4的神经细胞对淀粉样蛋白堆积更敏感,更容发生细胞凋亡和坏死。有趣的是,如果携带E4变异的人在运动中发生脑震荡或脑部受伤,他们会比携带正常E3型的人需要更长的时间才能恢复。
参考文献:
Bell RD, Sagare AP, Friedman AE, Zlokovic BV, et al. Transport pathways for clearance of human Alzheimer's amyloid beta-peptide and apolipoproteins E and J in the mouse central nervous system. J Cereb Blood Flow Metab. 2007 May;27(5):909-18. PMID: 17077814.
Boehm-Cagan A, Bar R, Liraz O, et al. ABCA1 Agonist Reverses the ApoE4-Driven Cognitive and Brain Pathologies. J Alzheimers Dis. 2016 Oct 4;54(3):1219-1233. PMID: 27567858.
Cohen SI, Linse S, Luheshi LM, et al. Proliferation of amyloid-β42 aggregates occurs through a secondary nucleation mechanism. Proc Natl Acad Sci U S A. 2013 Jun 11;110(24):9758-63. PMID: 23703910.
Frieden C, Wang H, Ho CMW. A mechanism for lipid binding to apoE and the role of intrinsically disordered regions coupled to domain-domain interactions. Proc Natl Acad Sci U S A. 2017 Jun 13;114(24):6292-6297.PMID: 28559318.
Mahley RW, Weisgraber KH, Huang Y. Apolipoprotein E: structure determines function, from atherosclerosis to Alzheimer's disease to AIDS. J Lipid Res. 2009 Apr;50 Suppl:S183-8. PMID: 19106071.
Mahley RW, Weisgraber KH, Huang Y. Apolipoprotein E4: a causative factor and therapeutic target in neuropathology, including Alzheimer's disease. Proc Natl Acad Sci U S A. 2006 Apr 11;103(15):5644-51. PMID: 16567625.
Mandrekar-Colucci S, Karlo JC, Landreth GE. Mechanisms underlying the rapid peroxisome proliferator-activated receptor-γ-mediated amyloid clearance and reversal of cognitive deficits in a murine model of Alzheimer's disease. J Neurosci. 2012 Jul 25;32(30):10117-28. PMID: 22836247.
Singh S, Trikha S, Bhowmick DC, Sarkar AA, Jeremic AMJ. Role of Cholesterol and Phospholipids in Amylin Misfolding, Aggregation and Etiology of Islet Amyloidosis. Adv Exp Med Biol. 2015; 855:95-116. PMID: 26149927.
Tai LM, Mehra S, Shete V, et al. Soluble apoE/Aβ complex: mechanism and therapeutic target for APOE4-induced AD risk. Mol Neurodegener. 2014 Jan 4;9:2. PMID: 24386905.
Wahrle SE, Jiang H, Parsadanian M, et al. ABCA1 is required for normal central nervous system ApoE levels and for lipidation of astrocyte-secreted apoE. J Neurosci. 2008 Nov 5;28(45):11445-53. PMID: 15269217.
Zhao J, Fu Y, Liu CC, et al. Retinoic acid isomers facilitate apolipoprotein E production and lipidation in astrocytes through the retinoid X receptor/retinoic acid receptor pathway. J Biol Chem. 2014 Apr 18;289(16):11282-92. PMID: 24599963.
Zhou H, Yang Z, Tian X, et al. Lanosterol Disrupts the Aggregation of Amyloid-β Peptides. ACS Chem Neurosci. 2019 Sep 18;10(9):4051-4060. PMID: 31369236.



合作咨询
![]() 肖女士
肖女士
![]() 021-33392297
021-33392297
![]() Kelly.Xiao@imsinoexpo.com
Kelly.Xiao@imsinoexpo.com
 2006-2025 上海博华国际展览有限公司版权所有(保留一切权利)
沪ICP备05034851号-57
2006-2025 上海博华国际展览有限公司版权所有(保留一切权利)
沪ICP备05034851号-57