在过去的十年中,靶向蛋白降解技术飞速发展,已成为一种新型的治疗策略。TPD技术可以利用细胞泛素依赖的蛋白水解系统有效降解目标蛋白。尽管到目前为止已经报道了多种PROTAC,但所使用的E3连接酶配体大多局限于CRBN或VHL配体。
此外,有研究表明基于CRBN配体或VHL配体的PROTACs会产生耐药性等问题,这些PROTACs由于E3连接酶的细胞类型特异性表达而显示出局限性。为了克服这些障碍,各种E3连接酶配体的开发已成为人们关注的焦点,以改进当前的PROTAC技术。
什么是PROTAC?
靶向蛋白降解(Targeted Protein Degradation, TPD)是一种新兴的治疗策略,被认为是克服传统药物开发局限性的解决方案。
TPD通过E3连接酶,进而诱导泛素蛋白酶体系统(UPS)选择性地降解目标蛋白(POI)。
PROTAC由一个与E3连接酶配体、一个靶向POI的配体和配体之间的化学连接链组成(图1)。与PROTAC一起,使用蛋白质标签的蛋白质降解策略已经开发出来,如dTAG、AiD和SMASh Tag技术,他们同样是通过诱导邻近的POI泛素化降解POI。
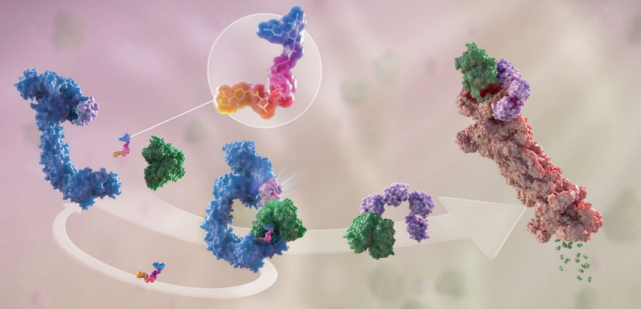
图1.PROTAC作用机制
图片来源:Arvinas官网
尽管人类细胞中有超过600种E3连接酶,但只有非常有限数量的E3连接酶被用于PROTAC技术(CRBN, VHL, IAP和MDM2)。目前为止,大多数PROTACs都局限于CRBN或VHL。
然而,最近关于CRBN或VHL的PROTACs耐药的研究强烈表明,要充分利用PROTAC策略,需要发现更多的E3连接酶配体。
此外,考虑到许多E3连接酶已被证明在特定类型的细胞或组织中过表达。例如,大脑(FBXL16, KCTD8)、胰 腺(ASB9)、骨骼肌(KLHL40, KLHL41)、睾丸(DCAF4L1)、输卵管(DCAF8L1)。
开发新的E3连接酶可能为PROTACs提供更好的机会,可以开发出具有更高的选择性和特异性的PROTAC,用于有效的疾病治疗。
已开发的E3泛素连接酶配体
CRBN配体
在20世纪50年代,沙利度胺(1)首先由celgene开发,作为孕妇晨吐的镇静剂。然而,在20世纪60年代初,沙利度胺因其严重的致畸作用被撤出市场。
而后,沙利度胺被重新开发为一种有前途的免疫调节亚胺药物(IMiD),并被批准用于治疗麻风结节性红斑(ENL)和多发性骨髓瘤(图2)。
然而,沙利度胺的作用机制一直未被阐明,直到2010年Ito等人发现沙利度胺的靶蛋白是E3泛素连接酶CRL4CRBN的一个亚基。除沙利度胺外,2011年,该团队也鉴定了泊马度胺(2)和来那度胺(3)的靶蛋白也为CRBN。
2018年,Crew小组报告了22种沙利度胺类似物,其中3种沙利度胺类似物(4,5,6)表现出更好的药理特性和良好的CRBN结合亲和力(4,5,6的Kd分别为555,549和111 nM)。他们发现,沙利度胺中邻苯二酰亚胺部分的化学修饰并没有导致CRBN结合亲和力的显著下降。
C4 Therapeutics申请了CRBN配体的合成专利,他们报道了各种哌啶2,6-二酮衍生物。荧光偏振(FP)实验表明化合物7是最 佳的CRBN配体之一。
此外,他们发现进一步简化得到的化合物8对CRBN同样具有纳摩尔的亲和力。
Hwang小组设计了1-氨基苯并三唑-戊二酰亚胺作为新的CRBN配体,并发现了TD-106(9),TD-106(9)为直接CRBN配体。
2019年,Kymera Therapeutics报告了各种CRBN配体,这些配体具有两个环,与不同的连接剂缀合。其中效果最好是化合物10和SB572027 (11)。目前,其中一个配体SB572027(11)被百济神州(Beigene)用于合成靶向BTK的PROTAC。
2021年,Rankovic小组用苯基取代了沙利度胺的邻苯二酰亚胺基团,合成了苯酰戊二酰亚胺(PG, 12)。与沙利度胺(t1/2 = 3.3 h)相比,PG (12, IC50 = 2.191μM)表现出更为出色的稳定性。
2021年,上海药物所王明亮团队申请了CRBN配体的合成专利,包括化合物13和14。
2021年,诺华公司开发了基于CRBN配体15 和16的BRD9降解剂。并且,这些降解剂可以在纳摩尔浓度可以有效降解BRD9 。
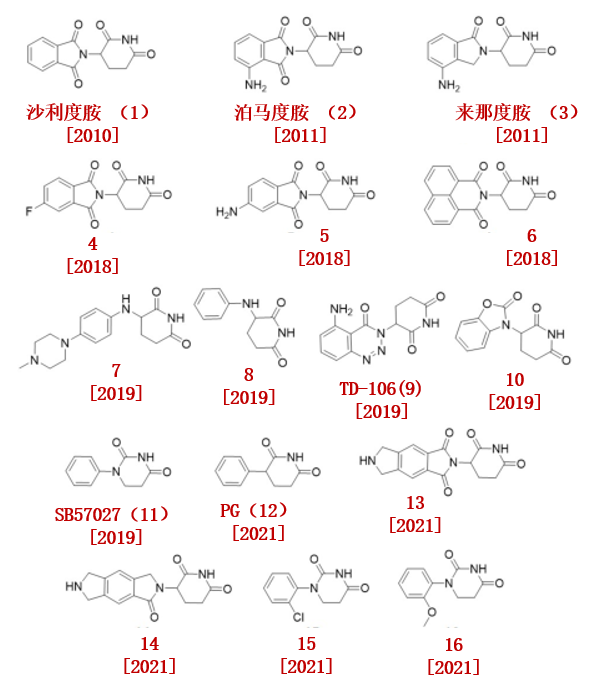
图2.CRBN配体化学结构
VHL配体
VHL蛋白是Cullin 2 E3连接酶的底物受体蛋白。缺氧诱导因子1α (HIF-1α)是VHL的底物蛋白之一。
2012年,Ciulli和Crews小组首次报道了一系列针对VHL的小分子抑制剂。他们合成了一个羟基脯氨酸衍生物库,发现了VHL/HIF-1α相互作用的抑制剂。其中,VHL配体17对VHL具有个位数的微摩尔活性。(图3)。
在后续研究中,Ciulli团队基于x射线晶体结构,设计优化了初始VHL配体,并于2014年报道了新的VHL配体VH03218(Kd = 185 nM)和19 (KD = 291 nM),具有纳摩尔结合亲和力。
2015年,Crews团队提出了HaloPROTAC的概念,同时设计出了VHL配体20,以此构建出的HaloPROTAC成功降解了目标蛋白 p38δ。
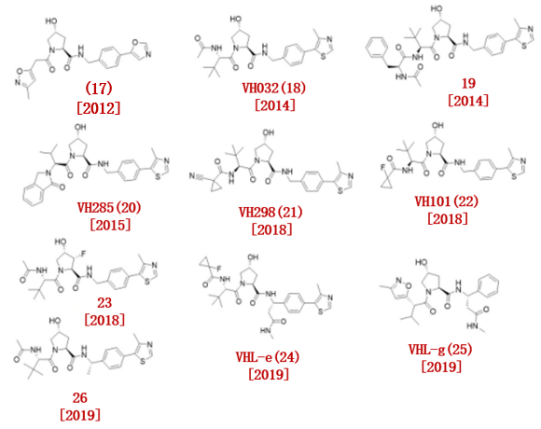
图3.已报道的VHL配体
2018年,Ciulli报告了VH298 (21, KD = 52 nM)和VH101 (22, KD = 16 nM),他们发现,增加VHL配体的亲脂性导致更高的细胞通透性和更高的VHL蛋白结合亲和力。
2018年,Ciulli团队报道了具有不同立体化学特征的氟羟脯氨酸(F-Hyp) VHL配体。他们合成了4种含有VHL配体的3-氟-4 -羟基脯氨酸的非对映异构体,发现VHL可以立体选择性地识别F-Hyp的(3R,4S)外映体(23)。
2019年,王少萌小组在前期工作的基础上,通过在VH101上引入(S)-甲基,发现了新的VHL配体。通过SAR研究,他们发现在(S)甲基上添加酰胺基团可以增加VHL配体的亲和力;基于FP的结合实验表明,VHL-e(24)与VHL具有较高的亲和力(IC50 = 190 nM)。
随后,王少萌团队进一步开发了弱结合VHL配体,VHL-g (25),同样对VHL蛋白显示出高度的亲和力。
Wang小组进一步开发了VHL配体(26),与 ER调节剂雷洛昔芬构建了靶向ER的高效降解剂。
IAP配体
凋亡蛋白抑制剂IAPs是细胞死亡的调节因子,可控制由多种刺激引起的凋亡过程。
2007年,Vucic小组开发了一种细胞凋亡抑制剂1和2 (c-IAP1和c-IAP2)拮抗剂(MV1, 27),它与IAP蛋白的IAP重复(BIR)结构域结合,导致 c-IAP的自泛素化和降解(KD = 5.8 nM)(图4)。
后来,Sekine等人报道了一种不同的cIAP1配体,贝斯他丁甲酯(ME-BS, 28)。
2012年,基因泰克公司发现了cIAP1/2、ML-IAP和XIAP的一种有效拮抗剂。他们通过基于晶体结构的SAR研究开发了一种广谱IAP抑制剂GDC-0152 (29)。
2012年,轩尼诗研究小组发现了一系列基于氨基哌啶的IAP抑制剂。他们发现,以船型固定的双环哌啶(30 (XIAP-BIR3的KD = 0.9 μ M)是cIAP1的有效抑制剂。
2013年,Cosford小组报道了一种有效的IAP拮抗剂。其中,IAP拮抗剂(31)对IAP的结合亲和力最好。
2014年,百时美施贵宝报道了异二聚IAP拮抗剂(32)对IAP具有高度亲和力,并具有出色的IAP抑制活性(IC50高达3.6 nM)。
此外。辉瑞基于化合物32合成了,一种BTK PROTACBC5P (DC50 = 182 nM)。
2017年,Naito团队开发了IAP配体LCL-161(33)及LCL-161衍生物(35、36),并开发了基于这些IAP配体的ER SNIPER, 成功降解了ER。
Astex制药公司通过基于片段的药物发现策略,成功发现了一种非拟肽cIAP1和XIAP抑制剂AT-IAP(34)。
AT-IAP(34)对XIAP和cIAP1表现出强烈的双重拮抗作用(XIAP EC50 = 5.1 nM, cIAP1 EC50 = 0.32 nM)。
2020年,葛兰素史克报告了一种基于哌柏西利的PROTAC,该PROTAC具有IAP 亲和力 (37),可用于降解CDK4和CDK6。
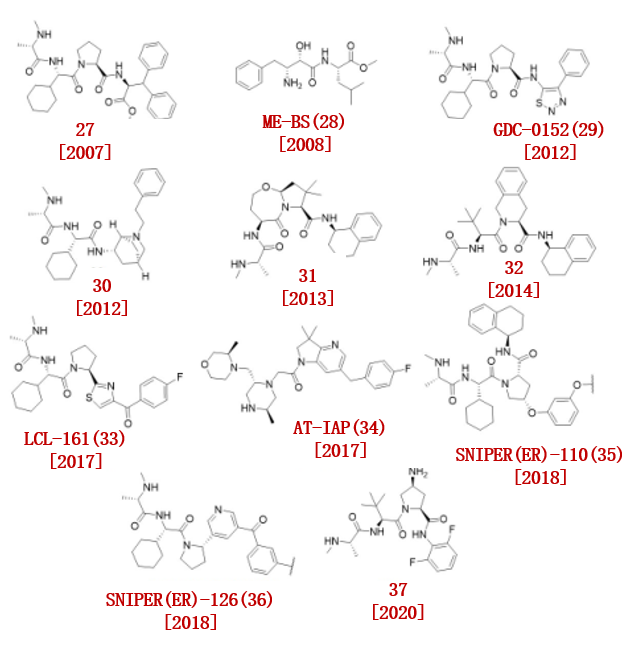
图4. 已报道的IAP配体
MDM2配体
MDM2蛋白是一种E3泛素连接酶,调节p53的泛素化和降解。
2004年,罗氏公司报告了一种强效的、高选择性的MDM2-p53相互作用的小分子抑制剂。他们通过筛选合成化合物库确定Nutlin3(38)对MDM2具有高度亲和力(图5)。
值得注意的是,顺式咪唑啉Nutlin-3(38)的两个对映体对MDM2具有高度不同的结合亲和力(对映体a = 13.6 μ M,对映体b = 0.09 μ M)。
2008年,Crews小组首次报道了基于MDM2的PROTAC,通过结合Nutlin-3(38)和一个具有PEG连接物的非甾体AR配体。
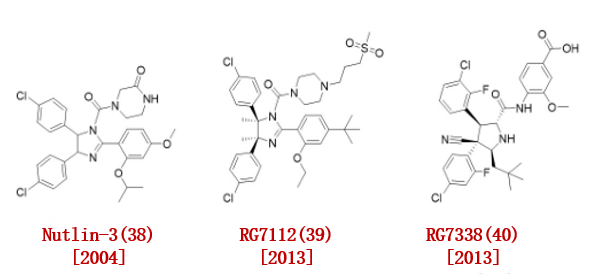
图5.已报道的MDM2配体
2013年,Roche根据p53-MDM2配合物的晶体结构对nutin -3(38)化合物进行优化,合成了一种新的MDM2抑制剂RG7112(39),在咪唑啉环上进行二甲基取代,用叔丁基取代甲氧基(IC50 = 18 nM)。RG7112是临床试验中首 个口服p53-MDM2抑制剂。
他们用吡咯烷部分取代了RG7112的咪唑啉结构,并引入了立体化学结构以获得更高的亲和力的MDM2抑制剂RG7388(40),具有优异的疗效和选择性 (IC50 = 6 nM)。
DCAF 配体
磺胺类衍生物因其抗菌、抗真菌、抗病毒和抗癌活性而备受关注。Nijhawan的研究报道,磺胺衍生物indisulam (41), E7820(42)和chloroquinoxaline sulfonamide (CQS, 43)作为一种分子胶,诱导E3连接酶和靶蛋白之间的蛋白-蛋白相互作用(图6)。
2019年,Cravatt团队使用化学蛋白质组学方法确定了DCAF16 E3连接酶作为他这些化合物(41、42、43、44)的靶蛋白。
基于类似的筛选策略,Cravatt小组也开发出E3连接酶DCAF11的亲电配体(45)。利用发现的配体45,他们合成了靶向AR PROTAC,并成功降解AR。
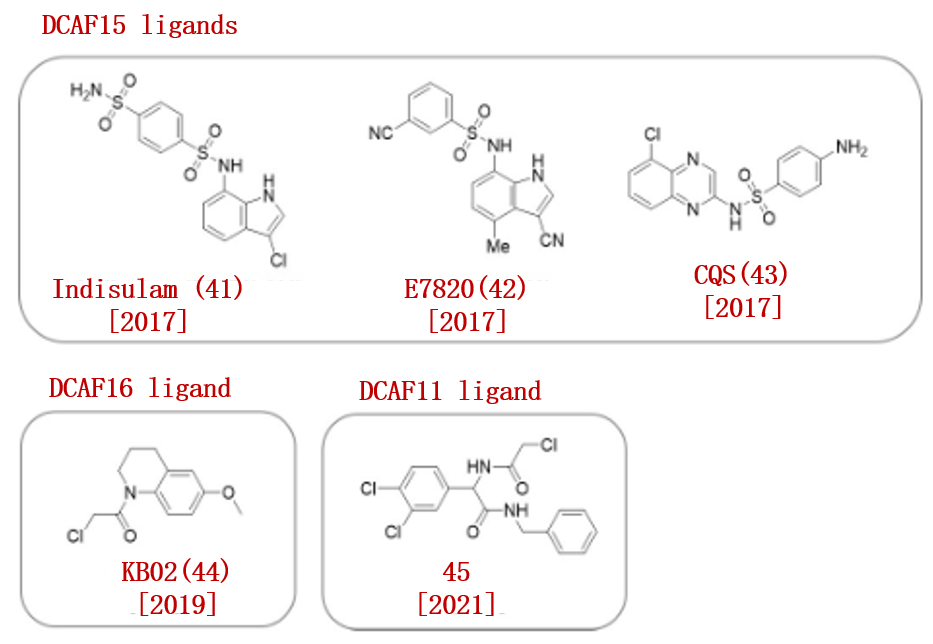
图6.已报道的DCAF配体
RNF配体
2019年,野村研究小组使用基于ABPP的共价配体筛选方法,报告了一组E3连接酶RNF4的配体。他们发现优化的共价配体CCW16(46)对RNF4具有高度亲和力。
此外,该小组还报道了Nimbolide(47),一种具有抗癌活性的天然产物,被鉴定为E3连接酶RNF114的共价配体。
在2021年,野村团队同样基于ABPP的方法发现了共价配体EN219(48),可以靶向RNF114 (IC50 = 470 nM)(图7)。
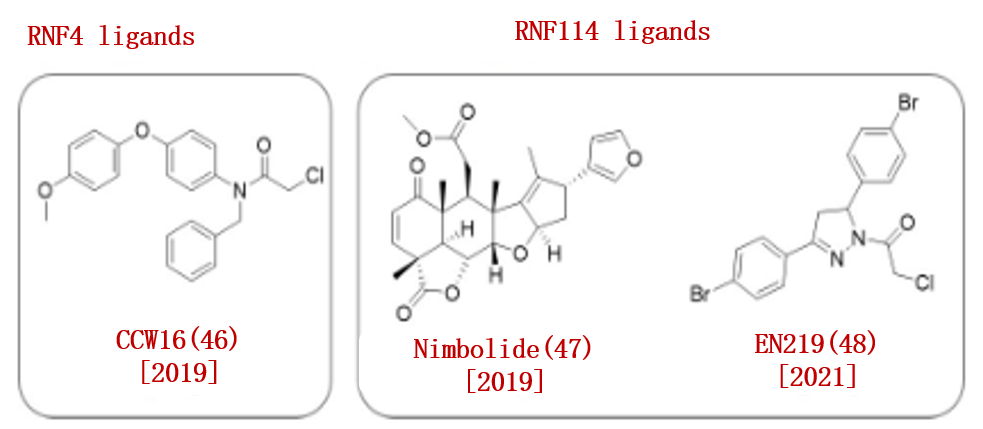
图7.已报道的RNF配体
AhR配体
2019年,Naito团队开发了一种新的PROTAC,可以招募芳基烃受体(AhR) E3连接酶复合物。他们将AhR配体(β-NF, 49)与ATRA结合,形成了一个嵌合分子β-NF-ATRA。β-NF-ATRA是一种招募CRABPs的PROTAC,通过泛素-蛋白酶体途径以AhR依赖的方式诱导CRABPI和CRABPII降解(图8)。
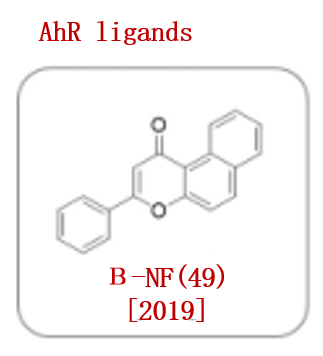
图8. 已报道的AhR配体
FEM1B配体
CUL2 E3连接酶FEM1B最近被发现是细胞对还原应激反应的重要调节因子。2022年,野村研究小组发现了一种基于氯乙酰胺的共价配体EN106(50)(图9),可作为一种FEM1B配体(IC50 = 2.2μM)。
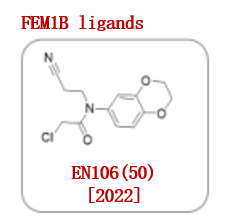
图9.已报道的FEM1B配体
KEAP1配体
研究表明KEAP1与Nrf2相互作用可以调节细胞保护蛋白。因此,基于KEAP1-Nrf2的蛋白-蛋白相互作用抑制剂的发现在应激相关疾病的治疗中引起了人们的关注。2020年,野村研究小组使用已知的KEAP1配体 (RTA 402,甲基巴多索隆(CDO- Me, 51)作为E3泛素连接酶配体,报告了一种可逆共价结合PROTAC。(图10)。
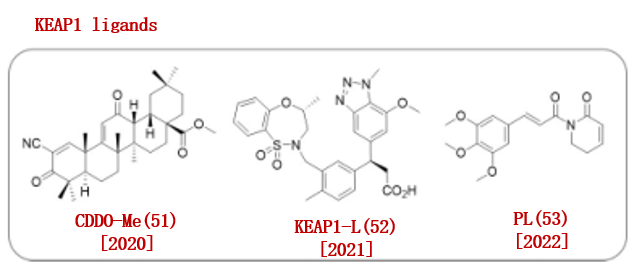
图10.已报道的KEAP1配体
2021年,金坚团队报道了E3连接酶KEAP152,并开发了一种招募KEAP1的PROTAC MS83。
2022年,Lv团队发现了一种天然产物Piperlongumine (PL, 53),作为E3连接酶配体。他们首先用竞争性的ABPP试验证实了PL(53)与多个E3连接酶结合。并且,他们发现当E3连接酶配体与靶蛋白配体偶联后,E3连接酶的选择性进一步提升。
小 结
PROTAC在过去的20年里已经发展成为治疗疾病领域的新策略。为了实现有效的TPD, E3连接酶配体和靶蛋白配体的选择是PROTAC设计的关键。尽管到目前为止,已经报道了多种E3泛素连接酶种类,但目前只有少数E3连接酶配体可用于TPD。
在过去的几十年里,PROTAC的大部分研究都集中在使用CRBN或VHL配体的各种药物靶蛋白的TPD上。然而,由于CRBN或VHL的细胞类型或组织类型依赖的表达差异化,靶蛋白降解经常受到抑制。此外,最近观察到对CRBN或VHL的PROTACs的耐药性。
而尚未开发的细胞或组织类型特异性E3连接酶将是一种新型PROTAC的研究基础,它可以以空间特异性的方式控制某种蛋白质的降解。因此,新型E3连接酶配体的发现将是一个重要的研究目标,以扩大PROTACs的应用前景。期待PROTAC技术的早日成熟,为患者带来福音!
参考文献
1.Discovery of E3 Ligase Ligands for T arget Protein Degradation. https://doi.org/10.3390/molecules27196515
2.Advances in targeted degradation of endogenous proteins. Cell Mol. Life Sci. 2019, 76,2761–2777
3.Targeted Protein Degradation Tools: Overview and Future Perspectives. Biology 2020, 9, 421
4.Major advances in targeted protein degradation: PROTACs, LYTACs, and MADTACs. J. Biol. Chem. 2021,296, 100647.
5.C. Molecular Glues for Targeted Protein Degradation: From Serendipity to Rational Discovery . J. Med. Chem. 2021, 64, 10606–10620.



合作咨询
![]() 肖女士
肖女士
![]() 021-33392297
021-33392297
![]() Kelly.Xiao@imsinoexpo.com
Kelly.Xiao@imsinoexpo.com
 2006-2025 上海博华国际展览有限公司版权所有(保留一切权利)
沪ICP备05034851号-57
2006-2025 上海博华国际展览有限公司版权所有(保留一切权利)
沪ICP备05034851号-57